Programme and abstracts as a pdf-file
|Bech|Brigham|Carey|Dawson|Denjean|Duchamp|Fyhn|Hohtola|Herbert|Körtner|Langseth|Maddocks|
|Marjoniemi|McKechnie|Nichelmann|Olson|Ophir|Pastukhov|Peltonen|Rashotte|
|Saarela|Schaub|Schleucher|Sedunova1|Sedunova2|Tzcshentke|
BODY COMPOSITION AND METABOLIC RATE IN YOUNG ALTRICIAL (EUROPEAN SHAG, PHALACROCORAX ARISTOTELIS) AND PRECOCIAL (PEKIN DUCK, ANAS PLATYRHYNCHOS) BIRDS
Bech C., Langseth, I. & Østnes, J.E.
Department of Zoology, Norwegian University of Science and Technology, N-7034 Trondheim, Norway
During the development of avian nestlings, their resting metabolic rate changes in a biphasic pattern with a peak value occurring approximately at the time of maximum growth. The maximal attained resting metabolic rate is often higher than that expected for an adult bird of similar body mass. In the present study we examined the possible influence of variations in the size of internal organs in "setting" the high resting and peak metabolic rate during development in a large altricial, the European shag (Phalacrocorax aristotelis), and a precocial (the Pekin duck, Anas platyrhynchos) species. In both species, resting metabolic rates (RMR, measured at thermoneutrality) and peak metabolic rates (PMR, cold-exposure induced) were measured at the age at which the RMR had reached its highest value during development. Thus, metabolic rates were measured in the ducklings at an age of 5 days, while the shag nestlings had their metabolic rates measured at an age of 15 days. Chicks of both species were sacrificed after the metabolic measurements, and their body composition were recorded.
In adult birds, the resting metabolic rate has been demonstrated to be correlated with the combined masses of the heart and kidney (Daan et al. 1989, 1990). However, for both the shag nestlings and the ducklings, there was no such correlation. In both species, however, there was a correlation between the RMR and the liver mass, and in the case of the shag nestlings, also between the RMR and the intestine length. In the ducklings, RMR in addition correlated with the sum of the internal organ masses. PMR correlated, in the ducklings, with the combined heart and kidney mass, while in the shag nestlings PMR correlated only with the intestine length. These results suggest that the internal constraints on metabolic rates may differ both between chicks and adults and also between chicks of different developmental mode. Thus, in chicks the 'supply organs' (e.g. intestine, liver) are apparently setting the pace of the development of the RMR, while in adults, the RMR is more a reflection of the activity of the 'consumer and distributing organs'. Also, the RMR of the altricial shag chicks seems to be more constrained by the development of the intestine compared to the RMR of the precocial ducklings.
Daan, S., Masman, D., Strijkstra, A. & Verhulst, S.
J.
Biol. Rhythms 4:267-283.(1989)
Daan, S., Masman, D. & Groenewold, A. Am. J. Physiol.
259:R333-R340 (1990)
USE OF TORPOR BY FREE-RANGING AUSTRALIAN OWLET-NIGHTJARS (AEGOTHELES CRISTATUS)
Brigham, R.M., Körtner, G. & Geiser, F.
Division of Zoology, University of New England, Armidale, Australia.
Current address of RMB: Biology, University of Regina, Regina, Canada.
With the exception of some data for Common Poorwills (Phaleanoptilus nuttallii) and anecdotal reports for a few other species, our knowledge about the use of torpor by free-ranging birds is very limited. In Australia, data on the temperature relations of birds is particularly scarce. Thus the purpose of our study was to assess the use of torpor by free-ranging Australian Owlet-nightjars which are unusual Caprimulgiformes in that they roost in tree hollows. We selected this species for study because of their relatively small body size (50 g), insect diet, nocturnal sedentary nature, taxonomic affiliation with other birds for whom the use of torpor is well documented and the cold winters (average July minimum Ta of -1 ºC ) in the study area. From data on poorwills (Brigham 1992), we generated the following prediction: owlet-nightjars would use daily torpor, especially on cold nights after a short bout of early evening foraging but, but only outside of the breeding season. We tracked 10 birds carrying temperature sensitive transmitters for a total of 842 days. Four different individuals entered torpor on a total of 76 days. Torpor bouts occurred predominantly during the daytime and only between 8 May and 8 September which is the cold season in the study area. The lowest skin temperature recorded for any bird was 19.6 ºC which is considerably higher than the 3 ºC temperature found in poorwills. Surprisingly, torpor was rarely used on winter nights. The birds typically foraged until dawn and when torpor was employed, aroused after bouts lasting only 1-3 hours. We speculate that the reasons for this could include: 1) owlet-nightjar's ability to forage all night using walking rather than just on the wing; 2) small size which limits fat deposition and or 3) tree roosting behaviour allowing birds to remain hidden in the daytime. On some occasions birds re-entered torpor in the afternoon (possibly they were warmed up by the sun in the morning and when the sun angle decreased, entered torpor again). As we predicted, torpor was not used during the breeding season, but this period also corresponds to the warm part of the year. Our results show that even though Australia is typically thought of as a very temperate continent, at least some of the avifauna employ torpor as a regular means of saving energy.
Brigham, R.M. 1992. Physiol. Zool. 65:457-472.
CAN BIRDS PREDICT THE ONSET AND SEVERITY OF SEVERE WINTER STORMS?
Carey, C. & Russell, C.
Dept. of Biology, University of Colorado, Boulder, Colorado USA
National Oceanic and Atmospheric Administration, Boulder, Colorado USA
The severity of winter storms varies with the depth of ice or snow cover, wind velocity, air temperatures, and the duration of the storm. These factors can pose significant challenges to birds wintering in cold climates by exacerbating thermoregulatory costs and restricting food availability to the point that typical defenses provided by winter acclimatization may not be sufficient for survival. Differential mortality during severe storms may have selected for the abilities to predict the onset and severity of winter storms and to make appropriate behavioral and/or physiological adjustments necessary for survival. However, little evidence exists that birds can predict snowstorms well in advance.
The questions addressed by this paper: what cues do winter storms give as they approach, how do birds sense these cues, and how accurate are they as predictors of the time of onset of precipitation and the severity (amount of precipitation and duration of the storm)? Barometric pressure, infrasound, wind speed and direction, cloudiness, and air ions are thought to be cues given by approaching storms. Of these, it is relatively certain that at least some species can sense changing wind direction, barometric pressure, and infrasound. An analysis of recent data gathered this last winter from a number of recording stations in California will be provided to indicate how reliable these cues are for predicting the amount and duration of rainstorms associated with an El Niño/Southern Oscillation event. Although these storms are not snowstorms, they will provide an index of the reliability of these cues for other types of storms.
GEOGRAPHIC AND CIRCUMSTANTIAL CORRELATES OF WINTER ACCLIMATIZATION IN SMALL BIRDS
Dawson, W. R.
Several small North American passerines with broad winter ranges show geographic variation in acclimatization state in this season. House finches (Carpodacus mexicanus) from areas having cold winters (Michigan, Colorado) increase cold resistance, whereas those from southern California, where winter conditions are milder, do not (Dawson et al., 1983; O'Connor, 1995). Additionally, dark-eyed juncos (Junco hyemalis) from South Dakota, where winter conditions can be severe, show greater cold resistance in winter than those from a milder area in Oregon (Swanson, 1993). Geographical variation in acclimatization state is also evident in extent of winter fattening. Californian house finches do not increase lipid accumulation at this season, unlike their conspecifics from colder areas (Dawson et al., 1983). Moreover, extent of winter fattening in dark-eyed juncos appears to vary inversely with temperature on a geographic basis (Rogers, 1995). Lipid mass also shows a significant correlation with lowest mean monthly temperature among populations of North American house sparrows, Passer domesticus (Blem, 1973). Studies of fat reserves of American goldfinches (Carduelis tristis) reveal a roughly similar pattern: southern Californian birds show no significant winter fattening, whereas those from eastern Washington State, Michigan, Wisconsin and New Jersey, where colder conditions prevail at this season, do (Dawson and Marsh, 1986; W. R. Dawson and J. Mansfield-Jones, unpublished data). The metabolic changes and associated lipid deposition comprising winter acclimatization of small passerines should impose substantial energetic costs. The geographic variation in acclimatization state noted above presumably serves to keep these costs commensurate with the extent of local climatic challenges, thereby avoiding expensive overcompensation. In evaluating this generalization, it is important to note certain exceptions, which will be discussed. Moreover, a tradeoff between the needs for matching energy reserves to the local situation and for minimizing predation risk, as well as a variety of other circumstances, can influence extent of winter fattening and thus acclimatization state within individual populations. To illustrate this situation, the possibility of effects on winter fattening of such things as the approach of winter storms and/or snowfall, food availability, social status, and incidence of predators will be reviewed. This indicates that consideration of behavioral ecology as well as of physiology and biochemistry is important for understanding winter acclimatization in small passerines.
Blem, C. R. Ornithol. Monogr. 14: 96-121 (1973).
Dawson, W. R., & Marsh, R. L. Physiol. Zool.
59: 357-368 (1986).
Dawson, W. R., Marsh, R. L., Buttemer, W. A., & Carey,
C. Physiol. Zool. 56: 353-369 (1983).
O'Connor, T. P. J. Comp. Physiol. B 165: 298-305
(1995).
Rogers, C. M. Physiol. Zool. 68: 277-289 (1995).
Swanson, D. L. J. Therm. Biol. 18: 275-281 (1993).
ARE THE MAMMALIAN-LIKE UNCOUPLING PROTEINS EXPRESSED IN BIRDS?
Denjean, F., Lachuer, J., Cohen-Adad, F., Georges, B., Barré, H. & Duchamp, C.
Laboratoire de Physiologie des Régulations Energétiques, Cellulaires et Moléculaires, UMR 5578 CNRS-Université Claude Bernard Lyon1, 43, Bld du 11 Nov. 1918, 69622 Villeurbanne, France.
Birds are well known to lack brown adipose tissue (BAT), the specialized thermogenic organ of small rodents. Further, no mammalian-like BAT uncoupling protein (UCP1), which plays a crucial role in BAT nonshivering thermogenesis (NST), was detected in avian adipose tissue (Saarela et al., 1991). Despite the lack of BAT, cold-acclimated Muscovy ducklings can develop NST (Barré et al., 1986a) mainly originating from skeletal muscle (Duchamp & Barré, 1993). Fatty acid-induced loose-coupling of mitochondrial respiration has been proposed as a potential mechanism for avian NST (Barré et al., 1986b), which could account for both the cold-induced NST and a lowering of the energetic efficiency of locomotion in vivo. In the absence of a mammalian-like UCP, the molecular mechanism of such uncoupling is not clear. It is conceivable that it is related to the expression of new members of the uncoupling protein familly, which have been recently described in mammals. Contrary to the BAT-specific UCP1, UCP2 is ubiquitously expressed in rat, mouse and human tissues (Fleury et al., 1997 ; Gimeno et al., 1997) and UCP3 is preferentially expressed in skeletal muscle and brown adipose tissue (Vidal-Puig et al., 1997). When transfected into yeast, these proteins partially uncouple mitochondrial respiration and could therefore play a role in controlling cellular energetic efficiency, renewing the interest in other tissues as potential sites of NST even in mammals. The question therefore arises as to whether UCPs similar to mammalian UCP2 and UCP3 are expressed in birds especially the species exhibiting NST in the cold and whether these proteins are involved in avian NST.
The expression of mammalian-like UCP mRNA has therefore been investigated
in cold-acclimated Muscovy ducklings by complementary molecular biology
techniques. Specific UCP1, UCP2 and UCP3 probes have been developped in
rats and used in Southern and Northern blots on duckling DNA and total
RNA. Reverse transcriptase and polymerase chain reaction (RT-PCR) assays
were developped using specific primers chosen from the published rat sequences
of the three UCPs. Degenerated primers selected in the more preserved regions
of the UCP sequences described in mammals and vegetables (Laloi et al.,
1997) have also been used in RT-PCR. Results obtained with these techniques
will be presented at the Symposium.
Barré, H., Cohen-Adad, F., Duchamp, C. & Rouanet,
J. L. J. Physiol. Lond. 375: 27-38 (1986a)
Barré, H., Nedergaard, J. & Cannon, B. Comp.
Biochem. Physiol. 85B: 346-348 (1986b)
Duchamp, C. & Barré, H. Am. J. Physiol.
265: R1076-R1083 (1993)
Fleury, C., Neverova, M., Collins, S., Raimbault, S.,
Champigny, O., Levi-Meyrueis, C., Bouillaud, F., Seldin, M. F., Surwit,
R. S., Ricquier, D. & Warden, C. H. Nat. Genet. 15: 269-272
(1997)
Gimeno, R. E., Dembski, M., Weng, X., Deng, N., Shyjan,
A. W., Gimeno, C. J., Iris, F., Ellis, S. J., Woolf, E. A. & Tartaglia,
L. A. Diabetes 46: 900-906 (1997)
Laloi, M., Klein, M., Riesmeier, J.W., Müller-Röber,
B., Fleury, C., Bouillaud, F. & Ricquier, D. Nature 389: 135-136
(1997)
Saarela, S., Keith, J. S., Hohtola, E. & Trayhurn,
P. Comp. Biochem. Physiol, 100B: 45-49 (1991)
Vidal-Puig, A., Solanes, G., Grujic, D., Flier, J. S.
& Lowell, B. B. Biochem. Biophys. Res. Commun. 235: 79-82 (1997)
REGULATORY, CELLULAR AND MOLECULAR ASPECTS OF AVIAN MUSCLE NST
Duchamp, C., Marmonier, F., Denjean, F., Lachuer, J., Eldershaw, T.P.D., Rouanet, J-L., Meister, G., Bénistant, C., Roussel, D., Cohen-Adad, F., Georges, B., & Barré, H.
Lab. Physiologie des Régulations Energétiques, Cellulaires et Moléculaires, UMR 5578 CNRS-Univ. Claude Bernard Lyon1, 43, Bld du 11 Nov. 1918, 69622 Villeurbanne, France.
Despite their lack of brown adipose tissue (BAT), the specialized thermogenic organ of rodents, a few species of birds including ducklings and penguins can develop nonshivering thermogenesis (NST) after prolonged exposure to cold (Barré et al., 1986; Duchamp et al., 1989). This non-BAT dependent cold-induced NST is primarily of skeletal muscle origin (Duchamp & Barré, 1993). Recent data on the regulatory, cellular and molecular mechanisms of avian muscle NST will be reviewed.
In vivo, NST development in 5-wk-old cold-acclimated (4°C, CA) muscovy ducklings is associated with a lowering of the apparent energetic efficiency of physical activity at thermoneutrality, further suggesting energetic alterations in skeletal muscles.
At the tissue level, CA-induced muscle NST is fueled by coordinated increases in fatty acid (FA) supply from adipose tissue, cellular uptake of lipoprotein-derived FA, and intracellular FA transport capacity. Indeed, basal and glucagon-induced lipolytic activities of fat fragments were higher in CA than in thermoneutral (25°C, TN) ducklings. Endothelial lipase activity per organ was much higher in red gastrocnemius muscle and liver of CA ducklings. The intracellular FA-binding capacity of small cytosolic proteins was also higher in gastrocnemius muscle and was linked to a higher content of an intracellular 15.4 kDa FA-binding protein (FABP). Mitochondrial carnitine-palmitoyl transferase activity was also increased. Similar results were obtained in cold-adapted king penguin chicks.
Perfused muscle preparations were developped to investigate the endocrine control of avian muscle NST activation. Results showed a higher resting muscle oxygen uptake (MO2) in CA than in TN ducklings. Further, MO2 was directly stimulated by catecholamines but not by glucagon. Higher in vitro thermogenic effects of catecholamines were observed in ducklings exhibiting muscle NST in vivo suggesting catecholamine involvement in muscle NST in vivo. The use of specific inhibitors showed that the higher resting MO2 in CA ducklings was related to a higher contribution of an uncoupled mitochondrial respiration while the thermogenic effect of catecholamines was linked to an ATP-coupled respiration.
At the subcellular level, two thermogenic processes have been implicated, one based on an uncoupling of mitochondrial oxidations and phosphorylations and the other on an increased ATP-hydrolysis by enhanced sarcoplasmic reticulum (SR) Ca2+-cycling. The molecular basis of the two processes have been investigated and FA or their derivatives have been involved in their control. Recent data showed that maximal palmitate-induced uncoupling does not abolish ATP production of skeletal muscle mitochondria, suggesting that mitochondrial and reticular thermogenic processes can sinergistically occur.
The control at the gene level of the functional adaptations of skeletal
muscle during cold acclimatation is currently under investigation and may
possibly involve the up-regulation of specific nuclear thyroid hormone
receptors.
Barré, H., Cohen-Adad, F., Duchamp, C. & Rouanet,
J. L. J. Physiol. Lond. 375: 27-38 (1986)
Duchamp, C. & Barré, H. Am. J. Physiol.
265: R1076-R1083 (1993)
Duchamp, C., Barré, H., Delage, D., Rouanet, J-L.,
Cohen-Adad, F. & Minaire, Y. Am. J. Physiol. 257: R744-R751
(1989)
VARIATION IN DAI LY ENERGY-EXP ENDITURE OF BLACK- LEGGED KITTIWAKES (RISSA TRIDACTYLA) DURING THE CHICK REARING PERIOD
Fyhn, M.1, Gabrielsen, G.W.2, Nordøy, E.S.1, Langseth, I.3 & Bech, C.3
1Department of Arctic Biology and Institute of Medical Biology, University of Tromsø, Norway.
2Norwegian Polar Institute, Tromsø, Norway.
3Department of Zoology, Norwegian University of Science and Technology,
Trondheim
In seabirds, the maximum parental energy demands occur during the chick rearing period (Drent & Daan 1980; Ricklefs 1983). A previous study on Black-legged Kittiwakes (Rissa tridactyla) has shown large individual variation of daily energy-expenditure (DEE) early in the chick rearing period (Gabrielsen et al. 1987). The purpose of the present study was to examine the variation in DEE both early (chicks 5-10 days old) and late (chicks 20-25 days old) in the chick rearing period and factors that determine this variation. DEE was estimated in free-ranging kittiwakes in a small colony at Svalbard (79°N, 10°E) using the doubly-labelled water method. Early in the chick rearing period, DEE was estimated in breeding individuals (n=20) of which DEE also was estimated in five birds late in the chick rearing period. Of the initial 20 birds used, 12 were captured at the end of the chick rearing period for measurements of resting metabolic rate (RMR) using open respirometry. Body mass was reduced by 9 % in females and 7 % in males between early and late chick rearing. Each parent attended the nest on average 12 h day-1 early and 5 h day-1 late in the chick rearing period. RMR ranged between 163 and 273 kJ day-1. Average RMR was 211 ± 30 (SD) kJ day-1 which is 33 % less than RMR found in incubating kittiwakes (Gabrielsen et al. 1987). DEE averaged 763 ± 96 (SD) kJ day-1 early and 930 ± 266 (SD) kJ day-1 late in the chick rearing period. This is 3.9 and 4.4 times RMR for the early and late chick rearing periods respectively. Individual variation in DEE could not be explained by variations in any of the following parameters: RMR, brood size (one or two chicks), sex, time spent off nest or chick age.
Drent, R.H. & Daan, S. Ardea 68:225-252 (1980) Gabrielsen, G.W., Mehlum, F. & Nagy, K.A. Condor 89:126-132 (3987) Ricklefs, R.E. Stud. Avian Biol. 8:84-94 (1983)
Hohtola, E.
Department of Biology, University of Oulu
90570 Oulu, Finland
The need to measure shivering in birds arises when the physiological basis for the cold-induced thermogenesis has to be verified. From a purely phenotypic viewpoint, indirect calorimetry (total thermogenesis) is enough to estimate the ability of a bird to fight metabolically against heat loss. In addition to being a relevant physiological question in its own right, the dissection of cold-induced thermogenesis to shivering and other putative components helps to clarify the phylogeny of endothermy.
Electromyography is presently the standard method for measuring the contractile activity of shivering muscles. Recording electromyograms (EMGs) from single muscle is relatively straightforward and the same electromyographic indices that are known to correlate with muscle force can be used. As the time average of EMG is zero, various parameters that measure the 'deviation' of the signal are used (Hohtola, 1982, Toien, 1992). These include the mean rectified or root mean square amplitude of the EMGs. These are equivalent to the statistical parameters known as mean deviation and standard deviation. In a number of avian species, such measures have been shown to correlate well and linearly with cold-induced whole-body thermogenesis measured by indirect calorimetry. In these species, various muscle groups start shivering at the same time and the shivering increases monotonically with decreasing cold load (e.g. Stevens et al., 1986). In birds, the pectoralis major is the major thermogenic organ for shivering in species that are agile flyers. Furthermore, it is electrically silent in a resting bird at thermoneutrality and, apart from flying, most forms of activity do not induce spurious electromyographic activity. This makes birds an attractive model for studying shivering thermogenesis.
Many studies have shown, however, that the onset temperature for shivering may vary between muscles (Aulie & Toien, 1988, Carey et al., 1989). Typically, highly aerobic muscles are recruited before more "white muscles". The onset temperatures can also change during ontogenesis as the muscle undergo changes in their metabolic machinery (Marjoniemi & Hohtola, unpubl.). In extreme cases shivering seems to be localized in a very restricted part of a large muscle group such as the iliotibialis in the leg muscles of the pheasant. Thus, the lack of shivering should be verified thoroughly before postulating other modes of heat production. Although various forms of vibrometry are not suitable for quantitative measurements of shivering, this method might be revived for detecting the presence or absence of shivering. This can be done as the vibrations induced by muscle tension can be measured even if the sensor is not positioned on the active muscle surface.
A further complication is that, even if EMG and actual thermogenesis show a linear relationship, this relationship may vary. In pigeons, the increment in total body thermogenesis for a unit increase in pectoral EMG is lower during the dark phase and is further lowered by fasting (Hohtola et al., in press).
Taken together, electromyographic measurements of shivering, albeit not very useful alone, produce, when combined with other thermoregulatory variables, important information on the function of one the main thermogenic effectors in homeotherms.
Aulie, A., and O. Tøien 1988. J. Comp. Physiol.
B 158:431-435.
Carey, C., R.M. Johnston, and A. Bekoff 1989. In: Thermal
Physiology 1989 (Mercer, J.B., ). Elsevier, Amsterdam, pp. 685-690.
Hohtola, E. 1982. Comp. Biochem. Physiol. A 73:159-166.
Hohtola, E., Henderson, R. & Rashotte, M. Amer.
J Physiol. in press.
Stevens, E.D., J. Ferguson, V.G. Thomas, and E. Hohtola
1986. Can. J. of Zool. 64:889-892.
Tøien, O. 1992. J. Thermal Biol. 17:357-366.
THE ONTOGENY OF ENDOTHERMIC REACTIONS IN THE PRECOCIAL DOMESTIC FOWL (GALLUS G. DOMESTICUS) - ALREADY AN EMBRYONIC PHENOMENON?
Herbert, S. & Prinzinger, R.
AK Stoffwechselphysiologie, Zoologisches Institut, Johann Wolfgang Goethe-Universität, Siesmayerstr. 70, D-60323 Frankfurt, Germany
Birds as well as mammals are homeothermic-endothermic organisms. They are able to actively regulate their body-temperature (Tb) on high levels even at low ambient temperatures (Ta). In contrast to poicilotherms they are independent of external warming: a decrease in Ta is compensated by an increase of the metabolic rate.
Precocial birds hatch highly developed: open eyes, well feathered, high mobility. Being independent of their parents they need -in contrast to altricial hatchlings- a functional thermoregulatory system immediately after they have left the egg. This leads to the question at which time the ability to regulate Tb occurs during the ontogeny. Is even the precocial embryo able to show endothermic reactions under cold load? We investigated this question in the domestic fowl (Gallus gallus domesticus).
Chicken eggs (12th to 21th day of incubation) were incubated at 37,5 ° C and 60 % rel. humidity. Every day in an experimental run the incubation-temperature was quickly lowered to 30 °C for 4 hours to test endothermic reactions. The oxygen consumption was measured in an open system (air flow rate: 20 L/h) with a MAGNOS 4G (Hartmann & Braun). Embryonic Tb (surface-temperature) was determined continuously by a thermosensor inserted under the eggshell (LUXTRON Flouroptic Thermometer, Model 790). After hatching the air flow rate was increased to 60 L/h; Tb was measured daily in the cloaca (Technotherm 9300, Testoterm).
Cooling the egg resulted in a progressive, typical poicilothermic decrease in metabolic rate and Tb throughout the whole incubation period. We observed no difference between young embryos and even pipped eggs. The embryos showed a clear resistance to this cooling: they withstood the decrease of Ta without any damage or delay in their development. They hatched at the time expected and synchronously with the controls incubated constantly under standard conditions (Ta = 37,5 °C)
The domestic fowl acquires its first ability of endothermic reaction immediately after hatching. Already 2 h-old chicks responded to Ta -decrease with a small but clear increase in heat production. Thermoregulatory ability improved rapidly and was relatively stable 2 days after hatching. In addition to this Tb of the young increased during the first days of life from 31,1 1,15 °C (directly after hatching) to 40,8 0,6 °C (5 th day of life).
Conclusion: It can be summarized that the transition from poicilothermy to homeothermy is an immediately post-hatching process. During the embryonic period there was absolutely no evidence of endothermic reaction.
THERMOREGULATORY BEHAVIOUR AND TORPOR OF FREE-RANGING AUSTRALIAN TAWNY FROGMOUTHS (Podargus strigoides)
Körtner, G. & Geiser, F.
Zoology, Biological Sciences, University of New England, Armidale 2351, Australia
Several small species (<100 g) of caprimulgiform birds are known to enter torpor. We wondered whether the much larger Australian Tawny Frogmouth (approx. 500 g) would also be heterothermic. We therefore investigated the thermoregulatory behaviour and temperature patterns in these birds under natural conditions in an open woodland close to Armidale (1000 m altitude) in New South Wales during winter. Climatic conditions during winter are characterised by cold nights, but mild temperatures during the day due to the predominantly dry and sunny weather.
Skin temperature (Tskin) of six Frogmouths was measured via external temperature sensitive radio transmitters attached to an elastic harness. One bird was implanted interperitoneally with a second transmitter. Temperature data were recorded automatically in 10 min intervals by custom made data-loggers. Air temperature (Ta) was measured at two locations by data-loggers in hourly intervals. Birds were radio-tracked daily and location, height and orientation of their day roost were determined.
Tskin correlated closely with body temperature (Tb), although Tskin was more influenced by Ta than in smaller mammals and birds in which external transmitters have been used to assess Tb. During the activity periods normothermic Tb were around 38 °C, but dropped to about 35.5 °C during rest. Between June and August, the three coldest months of the year, the Frogmouths frequently entered shallow torpor after a short activity period around dusk with Tskin falling to minimum values between 21-26 °C (Tb < 30 °C). The birds always rewarmed before sunrise to commence either a second short foraging period or to fly to one of their day roosts. The Ta threshold for entering shallow torpor appeared to be about 7 °C. On cold mornings Tskin would fall again around sunrise, but usually Tskin increased rapidly after this initial drop by passive heating through the sun. Consequently, a second prolonged torpor bout in the morning was observed only occasionally in one individual.
Frogmouths changed day-roosts regularly and selection of north-facing branches was significant, presumably to maximise sun-exposure. Thermoregulatory costs were further reduced by huddling of pairs, which occurred on 66.3% of all observations.
We conclude that Tawny Frogmouths like several related, but smaller, nightjars enter shallow torpor frequently under natural conditions when low Ta would demand high thermoregulatory costs for maintenance of normothermic Tb and reduce ground insect activity and thus food availability. The occurrence of torpor is mainly restricted to the night, the normal activity phase of Frogmouths, apparently because sun-exposed day-roosts and mild to warm climatic conditions during the day allow the maintenance of normothermic Tb at low energetic costs.
THE EARLY VENTILATORY RESPONSE TO COLD EXPOSURE IN PEKIN DUCKLINGS (ANAS PLATYRHYNCHOS)
Langseth, I. & Bech, C.
Department of Zoology, Norwegian University of Science and Technology, Trondheim, Norway
The development of thermoregulation in avian chicks has been the subject of several studies. However, very few studies have been focusing on the development of ventilatory parameters and how chicks accommodate their ventilation to different oxygen demands during cold exposure. In this study we measured metabolic rates, at thermoneutrality and during cold exposure, and the corresponding ventilatory parameters (respiratory frequency, f, and tidal volume, VT) in Pekin ducklings (Anas platyrhynchos) aged 0, 2, 5, 10 and 15 days. Calculations of minute volumes (VI) and lung oxygen extraction (Eo2) were based on these measurements.
The Pekin duck is a highly precocial species and even the hatchlings respond metabolically to a cold challenge. In newly hatched ducklings the peak metabolic rate (PMR) was 5.2 x the resting metabolic rate (RMR). The metabolic scope (PMR/RMR) decreased with age and was 2.8 in 15 days old ducklings. This was caused by an increase in RMR rather than by a decrease in PMR. There were also age related changes in the ventilatory parameters. During cold exposure ducklings of all age groups depended on increased ventilation rather than an increased Eo2 to accommodate their elevated oxygen demands. The tidal volume increased with age both at RMR and PMR. All the age-groups showed, during cold exposure, a constancy in their use of VT in regulating total ventilation, with a mean increase of 59 %. In contrast, f was relatively stable with age at RMR, with a mean of 30 breaths min-1. However, at PMR there was a much higher rise in f in newly hatched ducklings compared to the older ducklings. The increase in f was 229 % in hatchlings and 43 % in 15 day old ducklings.
The accommodation of the ventilation during cold exposure differ between the ducklings and adult Pekin ducks. In the ducklings increased oxygen consumption is mainly achieved by increased ventilation caused by changes in f. Adult ducks, on the other hand, mainly use increased Eo2 (Bech et al. 1984). However, even within the 15 first days of life there are pronounced changes in the ventilatory pattern in the ducklings, with a higher dependency of changes in respiration frequency immediately after hatching.
Bech, C., Johansen, K., Brent, R. & Nicol, S. Resp. Physiol. 57:103-112 (1984)
NOCTURNAL HYPOTHERMIA AND THERMOREGULATORY LIMITS OF AUSTRALIAN SILVEREYES (ZOSTEROPS LATERALIS)
Maddocks, T.A. & Geiser, F.
Division of Zoology, School of Biological Sciences, University of New England, Armidale, Australia
Little detailed information is available on the thermal physiology of small Australian birds, especially regarding their responses to extreme ambient temperatures (Ta). We therefore investigated the thermal responses and capabilities of an 11g Australian passerine, the silvereye (Zosterops lateralis), when exposed to a wide range of Ta, during both the photophase and scotophase. Silvereyes utilised nocturnal hypothermia on a daily basis, decreasing metabolic rate (MR) by up to 50 %, body temperature (Tb) on average by 3.5 °C, and thermal conductance by up to 30 % in comparison to resting photophase values. Within the thermoneutral zone (TNZ) during the scotophase (27.0-33.6 °C) the basal MR (BMR) was 2.30 ± 0.29 mL O2 g-1 h-1, which is 32 % lower than predicted, and Tb was 38.4 ± 0.8 °C. Below the TNZ, the maximum MR (HPmax) induced by helium-oxygen (79:21 %, He-O2) exposure was 15.64 ± 1.55 mL O2 g-1 h-1 at an effective lower limit Ta of -39.7 ± 6.1 °C during the photophase. During the scotophase the HPmax was 13.16 ± 1.58 mL O2 g-1 h-1 at an effective lower limt Ta of -41.7 ± 9.9 °C. The HPmax was sevenfold that of BMR, which is similar to that of similar sized passerines including arctic species. However, this high metabolic scope was primarily due to the low BMR of silvereyes. Above the TNZ, MR, Tb and conductance increased steeply and at Ta ~39 °C MR was about 1.7 times that of BMR, Tb was significantly elevated to 41.9 ± 0.8 °C and conductance was over 3 times that within the TNZ. Our study shows that silvereyes can withstand a Ta range of at least 81 °C, from about -42 to +39 °C. Moreover, they are able to produce heat that is sufficient to maintain a thermal difference between Tb and Ta of up to 80 °C. Their relatively low BMR, high TNZ and high capacity for heat production allows them to tolerate a wide Ta range and use of nocturnal hypothermia helps them to overcome periods of food shortages. These adaptations may explain to some extent why this species can live in a wide range of habitats and have such a wide distribution range.
SHIVERING THERMOGENESIS IN NESTLINGS OF DOMESTIC PIGEON (COLUMBA LIVIA)
Marjoniemi, K.and Hohtola, E.
Department of Biology, University of Oulu, Finland
Development of endothermy has been described in large number of both
precocial and altricial species, but there are only few accounts of ontogeny
of shivering in altricial nestlings, like Olson (1994). In this work we
studied the development of shivering thermogenesis in leg and breast muscles
of domestic pigeon.
Twenty pigeon nestlings (both sexes) aged 2, 4, 6, and 8 days were
used in experiments during August-September 1995. Nestlings were hatched
in outdoor aviaries in the Zoological Garden of the Department of Biology,
University of Oulu (65º N, 25º E), where they were incubated
and fed by their parents.
Shivering thermogenesis was studied by subjecting nestlings to decreasing
ambient temperature (0.35° C · min-1). To record shivering,
electromyograms were measured from leg and breast muscles (musculus
gastrocnemius and m. pectoralis, respectively) by bipolar subcutaneous
electrodes with simultaneous recordings of heat production (indirect calorimetry,
S-3A oxygen analyzer) and body temperature. The body temperature was measured
with a thermocouple inserted into the cloaca.
Median frequency of shivering was studied by analysing tape-recorded
EMG-samples with Data 6000 signal analyzer (Data Precision Inc.).
The amplitude of shivering was higher in pectorals. In cold, intensity
of EMG signal increased already at 2 days of age in both muscles studied
(P<0.05). However in gastrocnemius, EMG was caused by regular movements
of legs, and clear shivering begun only from the age of 4 days onwards.
Presumably, movements were due to behavioural thermoregulation (attempts
to huddle).
In pectorals of pigeons aged 2-6 days, steadily decreasing ambient
temperature did not cause a linear increase of EMG, but produced a sudden
onset to maximal shivering level. At the age of 8 days, shivering showed
linear increase also in pectoralis. The reason for abrupt onset might be
either poorly developed central control of shivering or a strategy to spare
energy as long as possible during cold exposure.
Shivering threshold temperature was higher in pectorals at all ages.
The decrease in threshold temperatures between days 2-4 is due to growth
of body mass. Increase in threshold temperatures between days 6-8 indicates
maturation of thermoregulation.
Shivering pattern changed from intermittent to continuos at the age
of 6 days both in pectorals and gastrocnemius. At this age nestlings showed
intensive heat production in cold. Between days 2-8, median frequency of
shivering increased in gastrocnemius (P<0.05) but did not change in
pectorals (P=0.274). Presumably, this indicates greater degree of maturation
of thermogenesis in leg muscles.
In conclusion, the maturation of muscular thermogenesis and thermoregulation
begins between 6-8 days of ages in pigeons. Breast muscles are the principal
site of thermogenesis, like in nestlings of red-winged blackbirds (Olson
1994). The beginning of intensive thermogenesis is synchronous with the
beginning of continuous shivering, which depends on the aerobic metabolism
of muscles (Hohtola and Stevens 1986).
Hohtola, E. and Stevens, E.D. 1986: The relationship of
muscle electrical activity, tremor and heat production to shivering thermogenesis
in Japanese quail. J. of Exp. Biol. 125:119-135.
Olson, J.M. 1994. The ontogeny of shivering thermogenesis
in the red-winged blackbird (Agelaius phoeniceus). J. of Exp. Biol.
191:59-88.
A HIGH THERMAL CONDUCTANCE FACILITATES EXOGENOUS HEAT ABSORPTION, BUT NECESSITATES OBLIGATORY HUDDLING IN THE WHITE-BACKED MOUSEBIRD, COLIUS COLIUS.
McKechnie, A.E. and Lovegrove, B.G.
The white-backed mousebird Colius colius is a frugivorous and folivorous member of the Coliiformes endemic to the arid regions of southern Africa. We investigated various aspects of this species' thermal physiology, with emphasis on the role of sunning behaviour and huddling.
Metabolic measurements of single birds and groups of up to six birds were made using standard open-system respirometry. Tb was measured using surgically-implanted temperature telemeters. Solar radiation was simulated in a constant environment chamber using a 650W light bulb.
The birds displayed conservative metabolic traits, a highly labile body temperature (Tb) , reduction in rest-phase energy expenditure by means of huddling behaviour, and the absorption of exogenous heat.
Basal metabolic rate was 40% lower than predicted, resulting in low rest-phase body temperatures (approx. 35C). The amplitude of circadian body temperature patterns was equivalent to 195% of the predicted value. Thermal conductance was higher than predicted. Comparisons of BMR, thermal conductance and Tb with published data using phylogenetically independent data sets (independent linear contrasts) revealed significant deviations from predicted values.
Heterothermy occurred during the rest phase at lower ambient temperatures. Periods of linear passive cooling were evident during the experimental scotophase, with cooling rates dependent on ambient temperature and group size. The lowest body temperature recorded was 26C, at an ambient temperature (Ta) of 5C.
Like other mousebird species investigated, C. colius conserves energy by means of huddling behaviour. In groups of six birds, rest-phase energy expenditure was reduced by 50% compared with single birds at Ta = 15C.
C. colius appears to make significant energy savings by absorbing solar radiation. At Ta = 15C, the birds reduced their energy expenditure by 20% over a one hour period when provided with simulated solar radiation.
A trade-off appears to exist in C. colius between the ability to maintain strict homeothermy, and the capacity to absorb exogenous heat. In short, a high thermal conductance facilitates heat absorption, but results in poor body temperature control in individuals during the rest phase. However, body temperature control is improved by huddling behaviour. This interplay of thermoregulatory variables suggests that huddling behaviour may be obligatory for the avoidance of nocturnal hypothermia. These traits have presumably evolved in response to selection pressures arising in an environment where food availability is spatially and temporally unpredictable.
THERMOREGULATORY HEAT PRODUCTION IN PRECOCIAL AVIAN EMBRYOS
Nichelmann, M.& Tzschentke, B.
Institut für Biologie der Humboldt-Universität zu Berlin, AG Perinatale Anpassung, Invalidenstraße 43, 10155 Berlin, Germany
It is well known that newly hatched precocial birds increase the thermoregulatory heat production (HP) in a cold environment whereas embryos of these species, also immediately before hatching, have never an endothermic reaction (Nichelmann et al., 1998). It has been shown that Muscovy duck embryos aged between 19 and 34 days decreased the oxygen consumption between 37.5°C and 34.0°C ambient temperature (Ta) and between 37.5 and 39.5°C Ta after an 100-min-temperature influence, so that the summit metabolism is situated at normal incubation temperature. The thermal neutral temperature is obtained at 40.5°C until day 25 of incubation, then it decreased to 39.0°C.
Methods
Because a 100-min-influence of low or high Ta may be connected with an hypothermia or hyperthermia in avian embryos, respectively, experiments in chicken (Gallus gallus) and Muscovy duck (Cairina moschata) embryos were carried out in which the eggs were inserted in a temperated chamber and the oxygen consumption and the body temperature (colonic temperature, Tc, and temperature of the allantoic fluid ,Taf) were measured continuously (Holland et al. 1998).
In other experiments, Muscovy duck embryos were incubated between the 23rd and 27th day at Ta of 34.5°C and HP was estimated in comparison to birds incubated at 37.5°C.
Results and Discussion
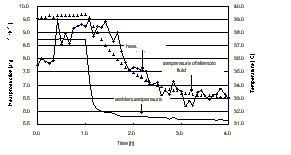
Fig. 1: Heat production, temperature of the allantoic fluid after decrease of ambient temperature in a Muscovy duck embryo (34th day of incubation).
Fig. 2: Heat production at different ambient temperatures in different incubated Muscovy duck embryos (27th day of incubation)
1. A decrease of Ta in Muscovy duck embryos as well as in chicken embryos usually resulted in a decrease of Taf, Tc and HP. Only in some embryos at the last day of incubation, shortly before hatching, a small increase in HP was obtained. It may be due to the post-plateau-phase increase or due to a net increase of thermoregulatory HP.
2. Muscovy duck embryos incubated at 34.5°C had at Ta of 36°C a higher HP than normal incubated embryos.
3. The measured HP of avian embryos is the result of the depressing influence of the Q10-effect and the stimulatory influence of the central nervous generated thermoregulatory HP. Because of this, a Q10-value of more than 2.0 demonstrates the absence of endothermy (Nichelmann et al., 1998). Between 39.0 and 37.5°C Taf a 34-day old Muscovy duck embryo shows a net increase in HP during the cooling procedure, it is also endothermic between 37.5 and 35.7°C but shows a net decrease in HP, and endothermy disappeared at Taf lower than 35.7°C. Generally, the older the embryo, the lower the minimal Q10 obtained, the lower the threshold temperature for a Q10 of 2.0 and the higher the Taf at the minimal Taf. The results suggest that the endothermic reaction in Muscovy duck occurs very early during the embryonic development and improve further until hatching.
Obviously, prenatal temperature experiences stimulate the thermoregulatory HP of embryos.
Holland, S.; Höchel, J.; Burmeister, A.; Janke, O.;
Nichelmann, M. J. therm.Biol. in press: 1998
Nichelmann, M.; Burmeister, A.; Janke, O.; Höchel,
J.; Tzschentke, B. J. therm. Biol. in press: 1998
ONTOGENETIC CHANGES IN CATABOLIC ENZYME PROFILE IN SKELETAL MUSCLES OF TWO SPECIES OF SHOREBIRD, THE DUNLIN AND WHIMBREL
Olson, J.M., Krijgsveld, K.L.1, and Ricklefs, R.E.2
Department of Biology, Villanova University, Villanova, PA. 1University of Groningen, 2University of Missouri.
Shorebird species nesting at high latitudes are exposed to variable
temperatures during the breeding season. Although the development of young
is usually classified as precocial or semiprecocial, chicks often experience
significant improvements in thermoregulatory capability during the post-hatching
period. For example, previous work on dunlin and whimbrel nesting near
Churchill, Manitoba (Canada) found that young chicks are capable of only
modest increases in metabolic rate in response to gradually decreasing
ambient temperatures (Williams, Ricklefs, and Visser, in prep.). Because
of incomplete development of thermoregulation, newly hatched and young
chicks of many species are more susceptible to hypothermia, and are therefore
especially vulnerable to the deleterious effects of low ambient temperatures.
Because chicks leave the nest soon after hatching, and must maintain locomotor
capabilities for both foraging and escape from predators, it is reasonable
to hypothesize that skeletal muscles involved in shivering thermogenesis
and/or locomotion will develop quickly and be less sensitive to lower body
temperatures, especially in younger chicks. We also hypothesized that muscles
will develop at different rates in a small (dunlin, Calidris alpina;
~4g at hatch) and large (whimbrel, Numenius phaeopus; ~20g at hatch)
shorebird. To test these hypotheses, we measured the activities of three
catabolic enzymes in dunlin and whimbrel to assess the capability of the
pectoralis muscle and mixed leg musculature to produce ATP. Specifically,
we assayed citrate synthase (CS; used as an indicator of aerobic capacity),
pyruvate kinase (PK; indicator of glycolytic capacity), and b-hydroxyacyl-dehydrogenase
(HOAD; indicator of the capacity for oxidation of fatty acids). Chicks
of dunlin and whimbrel were hand-raised in the lab in Churchill. Pectoralis
and mixed leg muscle samples were dissected from 1, 2, 4, 8, 12, and 16d
old chicks (n=2 at each age). In addition, muscle samples were collected
from 2 adults of each species. To assess the temperature sensitivity of
the catabolic enzymes, we measured activities at five temperatures between
17.5° and 43.0°C, a range which includes body temperatures potentially
experienced by the chicks. Generally, the activities of the three enzymes
(expressed per g fresh mass) in the pectoralis muscle increased ontogenetically,
with the highest activities recorded in the adults. Activities in the leg,
however, were often highest in chicks <16d old. In addition,
enzymatic activity is generally higher in smaller dunlin chicks than in
whimbrel at the corresponding age. No strong evidence exists, however,
that muscles of younger chicks possess catabolic enzymes with a lower thermal
sensitivity. The earlier biochemical development of muscles in dunlin is
consistent with the prediction that smaller animals with higher S:V ratios
and consequently higher heat flux need higher heat-generating capabilities.
That enzyme activity is sensitive to temperature in young chicks with large
S:V ratios and generally less well-developed thermoregulatory capabilities
underscores the importance of periodic brooding by the adult.
CUTANEOUS BLOOD FLOW IN THE PIGEON: A CRUCIAL COMPONENT IN CUTANEOUS WATER EVAPORATION?
Ophir, E.1,2, Arieli,Y.1,2,Horowitz, M.2 and Marder, J.1
1 Department of Cell and Animal Biology, Life Sciences, 2 Department of Physiology, Hadassah Medical School, The Hebrew University, Jerusalem, Israel
Previous studies strongly suggest that the heat-acclimated pigeon uses
cutaneous water evaporation (CWE) as the preferred route for heat dissipation
(ref. 1). It was also shown that the principal trait that distinguishes
heat acclimated pigeons (HAs) from non-acclimated (NAs) or cold-acclimated
(CAs) ones is the presence of controlled heat-induced CWE. This characteristic
evidently enables the pigeon to sustain full annual cycle at a daily exposure
to extremely high temperatures(Ta=60°c). The control mechanism involves
regulation of b-adrenergic (negative effect) and a2-adrenergic (positive
effect) stimulation(ref. 2). We substantiated that micro-structural adjustments
in the capillary wall are coupled to CWE (ref. 3). However, solid evidence
for the physiological events that take place in this phenomenon is not
yet available. In the present study we tested the hypothesis that changes
in cutaneous blood flow are a crucial component of this adaptation. Methods:
Cutaneous blood flow was measured by: 1) ultrasonic flowmetry (USF), for
single vessels and 2) laser Doppler flowmetry (LDF), for tissue perfusion.
Blood flow, concomitantly with CWE and rectal and skin temperatures were
measured under heat exposure (Ta=48°C)or following propranolol (P,
1.5 mg/kg, i.m.) or clonidine (C, 0.1mg/kg, i.m.) administration. Results:
Both P and C induced a significant increase in arterial blood flow in HAs
(2.7 and 2.2 fold, respectively)while venous blood flow showed a significant
decrease (0.4 and 0.5 fold, respectively). No significant changes in arterial
or venous blood flow were found in NAs or CAs following similar treatments.
By using LDF we found a significant increase (1.33 fold) in total blood
perfusion in the dorsal skin of HAs following P administration. In contrast,
a significant decrease (0.73 fold) was found in NAs. Injection of C resulted
in a significant decrease of total blood perfusion in dorsal skin of both
HAs or NAs (0.42 and 0.46 fold, respectively). Heat exposure increased
blood perfusion in both HAs and NAs (2.5 and 1.8 fold, respectively). Conclusions:
As shown by C effect, augmentation in cutaneous blood perfusion is probably
not a prerequisite for CWE. Essential differences in parallel arterial
and venous blood flow suggest that: 1) A particular fraction of the blood
volume entering the skin leaks out of the capillaries into the interstitial
space. This route may therefore cause tissue hydration. In this case, a
possible complex of lymph network may thus be responsible for the control
of the removal of excess water. 2) At least part of the vasomotor control
may be executed at the venous side. If this is the case, it is reasonable
to assume that capillary pressure is increased not solely by negative arterial
vasomotor response, but also by resistance augmentation somewhere along
the venous tree, leading to augmented A-V pressure difference. This pressure
rise may serve as an important driving force for the water extravasation
observed in the heat-exposed HAs.
1. Marder, J. and Gavrieli-Levin, I., J. Appl. Physiol.
62:952-958 (1987)
2. Ophir, E., et al., Eur. J. Physiol. Suppl. 430:R214
(1995)
3. Arieli, Y. et al. Am. J. Physiol. (1998) (submitted)
INTERACTIONS BETWEEN BODY TEMPERATURES AND SLOW WAVE SLEEP IN FED AND FASTED PIGEONS
Pastukhov, Iu.F.1, Rashotte, M.E.2, Poliakov, E.L.1, Sedunova,E.V.1, Henderson, R.P.2,Yekimova, I.V.1, Guselnikova, E.A.1 & Zimin, A.L.1
1Sechenov Institute of Evolutionary Physiology & Biochemistry, Russian Academy of Sciences,
St. Petersburg, Russia; 2 Department of Psychology, Florida State University, Tallahassee,
Florida, USA
In mammals, decreased metabolism, core and brain temperatures (Tb and Tbr) and increased total time spent in slow wave sleep (SWS TT) are considered to be interconnected homeostatic phenomena (Heller & Glotzbach, 1977; McGinty & Szymusiak, 1990). Less information is available on interrelations between Tb/Tbr and SWS in birds whose thermoregulation and sleep characteristics are substantially differ from those in mammals. To provide more data on this question, three series of studies were carried out in pigeons Columba livia.
1. In fed pigeons, nocturnal Tb decreased by 1.2 °C and SWS TT increased by 35% during the dark phase in comparison with light phase. In fasted pigeons (20% reduction of body weight), nocturnal Tb decreased by 3.1 °C and SWS TT was 91% greater than in the light phase. No differences in paradoxical sleep (PS) TT or in transitional states (TS) TT [similar to drowsiness] were found between fed and fasted pigeons in the dark phase. The fasted pigeons showed reduced PS TT in the light phase, however, and individual pigeons with low light-phase Tb had had greater SWS TT in that phase. Thus, nocturnal hypothermia in pigeons fasted to a weight loss of 20% is associated with SWS TT increase.
2. Two types of SWS episodes are found during the dark phase in fed and fasted pigeons, and these can be distinguished by accompanying changes in Tbr: in Type A, Tbr decreases by 0.05 to 0.10 °C; in Type B, the change in Tbr is less pronounced. We have identified two sequential sub-stages in Type A episodes: in A1 there is a decrease in Tbr; in A2 Tbr remains at a plateau or shows a small increase. The decrease in Tbr during A1is closely connected with A1 duration; the Tbr increase during A2 positively correlates with TS TT in that sub-stage. The proportion of time spent in TS during A2 is twice as great as in A1. The nature of the changes in Tbr during SWS in fasted pigeons compared to fed ones, might be connected with prolongation of episodes and SWS TT in the fasted birds. We propose that the often-reported decrease in Tbr during SWS is characteristic of only some SWS episodes, and only for the first sub-stage of such episodes.
Heller, H.C. & Glotzbach, S.F. Int. Rev. Physiol.
15:147-188 (1977)
McGinty, D. & Szymusiak, R. Trends Neurosci. 13:
480-487 (1990)
ROLE OF THE SKIN IN AVIAN THERMOREGULATION
Peltonen, L.1,4 , Arieli, Y.2, Pyörnilä, A.1 & Marder, J. 3
1Department of Biology, Zoology, University of Oulu, FIN-90571 Oulu
2Naval Medical Institute, Technion Israel, Haifa, Israel
3Department of Cell and Animal Biology, Laboratory of Environmental Physiology, Hebrew University of Jerusalem, Israel
4Present address: Department of Basic Veterinary Medicine, Physiology, 00014 University of Helsinki, Finland
The skin as a site for evaporative heat loss was studied in cold and heat-acclimated (CAC & HAC) pigeons (Columba livia). The structure of the skin was assessed by transmission electron microscopy and cutaneous water evaporation (CWE) by measuring the skin resistance to water vapour diffusion with a diffusive resistance meter (see Marder 1983; Marder & Ben-Asher 1983).
CWE in CAC pigeons is minimal, as indicated by high values for skin resistance (>50.0 s/m). The lack of any microvasculature adjacent to the epidermis indicates that the blood flow is shunted away from the periphery. This maximizes heat storage and minimizes cutaneous heat loss. The corneocytes in the stratum corneum (SC) are thin and they lack abundance of keratin bundles within the matrix originating from the keratohyalin granules of the viable epidermis. The whole viable epidermis from the stratum basale to the stratum transitivum (ST) is characterized by a large number of free lipid droplets and large multigranular bodies (MGB), but only a few, small keratohyalin granules. The formation of a mammalian-type lamellar water barrier in the extracellular spaces of the SC points to secretion of MGBs at the ST-SC interface. Such a barrier has been previously described in birds only in severely dehydrated individuals (Menon et al. 1989, 1996) and nestlings (Menon et al. 1987).
HAC pigeons have low skin resistance (0.0-3.50 s/m), and they use CWE exclusively under conditions of very high ambient temperature and low relative humidity. There are patchy, modified areas in the epidermis and the underlying vascular bed where water permeability is increased and the barrier forming machinery in the upper viable layers, the stratum transitivum and the stratum intermediale (SI) has deteriorated. While the epidermal cells are swollen and depleted in lipids, the keratohyalin granules are larger and more numerous than in the CAC birds. The superficial dermis contains a rich microvasculature with simple capillaries and flattened postcapillary venules.
The results show that the epidermis of HAC pigeons differs markedly from that of CAC birds, and that this difference is associated with the degree of cutaneous heat loss. In cold this heat loss is minimized and a tight, lamellar water barrier is formed, but in the heat modified skin patches are formed indicating increased permeability to water. Even though avians lack sudoriferous glands, the present results point to a sweating-like secretory function of specialized passages formed by changes in the vascular tree underlying certain epidermal areas.
In conclusion, the avian skin can have both an evaporative/secretory
and a barrier role in temperature regulation. The relatively loose skin
structure of non-challenged pigeons (Menon et al. 1986) can be modified,
directly or indirectly, by environmental factors in relation to the need
for cutaneous evaporative heat loss.
Marder, J. Comp. Biochem. Physiol. 75A: 433-439
(1983)
Marder, J. & Ben-Asher, J. Comp. Biochem. Physiol.
75A: 425-431 (1983)
Menon, G.K., Brown, B. & Elias, P.M. Tissue and
Cell 18: 71-82 (1986)
Menon, G.K., Baptista, L.F., Elias, P.M. & Bouvier,
M. Ibis 130: 503-511 (1987)
Menon, G.K., Baptista, L.F., Brown, B.E. & Elias,
P. M. Tissue and Cell 21: 83-92 (1989)
Menon, G.K., Maderson, P.F.A., Drewes, R.C., Baptista,
L.F., Price, L.F. & Elias, P.M. J. Morphol. 227: 1-13 (1996)
CIRCADIAN VARIATION IN THE PIGEON'S BODY TEMPERATURE:
ROLE OF DIGESTION, SHIVERING AND LOCOMOTOR ACTIVITY
Rashotte, M.E. & Chambers, J.B.
Department of Psychology, Florida State University, Tallahassee USA
Daily food intake has a pronounced influence on the pigeon's dark-phase Tb. In Ad Lib Feeding, nocturnal Tb is 1 to 1.5 °C lower than in the daytime. During several days of Fasting there is progressively greater nocturnal hypothermia with day-night differences ultimately reaching 7 to 8 °C. In Restricted Feeding it is possible to maintain a fixed level of nocturnal hypothermia by providing a constant volume of food each day. We utilized these three feeding conditions to investigate the thermogenic effects of digestion, shivering and locomotor activity on the circadian cycle of Tb.
1. Fasting. In the dark phase, the relationship between Tb and pectoral shivering can be studied without influence from locomotor activity or food-digestion which do not occur in that phase in fasted pigeons. When Ta = 21 °C, shivering: a) is suppressed near the beginning of the dark phase while the nocturnal fall in Tb is occurring, b) is increased while a hypothermic plateau in Tb is maintained, and c) is greatly enhanced when Tb rises before lights-on. Across the hours of the dark phase, individual pigeons show striking differences in the time course of changes in Tb (and in the associated shivering response). When Ta = 28 °C, fasting results in a similar pattern of nocturnal hypothermia, but without shivering. The involvement of non-shivering thermogenesis (NST) is suggested, particularly because Tb rises several degrees near the end of the dark phase without evidence of shivering or locomotor activity.
In the light phase, it is possible to observe interactions between shivering and locomotor activity without the influence of digestion because fasted pigeons show high levels of locomotor activity. The data indicate that shivering and locomotor activity combine in a complimentary fashion to influence Tb.
2. Restricted Feeding. In the dark phase, Tb falls to a nocturnal plateau that is directly related to the amount of food consumed late in the preceding light phase. The impact of different levels of food intake (and digestive activity) on the relationship between nocturnal Tb and shivering was investigated in this procedure. Shivering and digestion seem to combine in a complimentary fashion: as the amount of food consumed increases, the plateau of dark-phase Tb is elevated and dark-phase shivering is suppressed. Locomotor activity is not a factor in this result.
An interesting finding is that consumption of a given volume of non-nutritive pellets results in a higher nocturnal plateau in Tb than does consumption of the same volume of food pellets. This result seems related to elevated shivering during the night after non-nutritive pellets are consumed.
To investigate possible changes in the threshold for shivering at different
points in the dark phase, we imposed acute cold ambient temperatures. A
lower Ta was needed to initiate shivering at the beginning of the dark
phase than in the mid- or late- parts of the dark phase.
Geran, L.C. & Rashotte, M.E. Naturwissenschaften
84:
350-353 (1997)
Hohtola, E., Henderson, R.P. & Rashotte, M.E. Am.
J. Physiol. (in press)
MELATONIN, A CANDIDATE OF SIGNALLING MOLECULE FOR ENERGY SAVING
Saarela, S.1, Vuori, M.1, Eloranta, E.2 & Vakkuri, O.2
1Department of Biology and 2Department of Physiology, University of Oulu, Finland
Homeotherms including birds living at high latitudes encounter often simultaneously extremes of cold ambient temperature (Ta), darkness and food scarcity. Survival under these conditions requires the controlled integration of photic, thermal and energy balance information leading to optimal adjustments of multiple effectors from the level of behaviour and thermoregulation down to cellular metabolism. The interaction between thermoregulation and feeding behaviour is particularly apparent during the inactive phase of circadian cycle when dynamic changes occur in body temperature (Tb). Diurnal animals display a circadian rhythm of body temperature (Tb) exhibiting high Tb during the day and low at night. The magnitude of Tb changes is related to the animal's energy reserves. The amount of food ingested during the day seems to follow the adjustment of metabolism and body temperature to the proper level (Hohtola et al 1991, Rashotte et al 1993). Necessity for energy saving is followed by inadequacy of food ingestion. The question arises whether there is a chemical signal molecule that transmits energy balance information to multiple effector mechanisms of thermoregulation and behaviour. We study here the option that melatonin synthesised in digestive system of the quail may act as a signalling molecule for energy saving. This idea was hypothesised first by Saarela and Reiter (1993). Melatonin is one appropriate candidate since it drives circadian and seasonal rhythms both in birds and mammals. Secondly plasma melatonin rhythm exhibits a mirror profile for Tb of diurnal animals including a man. Moreover fasting of mice increases melatonin concentration in intestine (Bubenik et al. 1992).
Quails lived under 12:12 LD photoperiod exhibit significantly higher melatonin concentration both in plasma and digestive system in the midnight than at noon. The difference was 76x in plasma, 13.3x in the small intestine, 4.7x in the large intestine and 7.1x in the stomach. Melatonin concentration of digestive system was higher in 72 h fasted quails than in controls both at noon and at night. In the small intestine day values were 338 pg/g vs. 137 pg/g, and night values 6 747 pg/g vs. 1 816 pg/g, respectively. Similarly melatonin concentrations of colon and stomach and plasma were significantly higher in fasted quails than in controls. According to the correlation analysis changes in plasma melatonin concentrations due to pineal and retinal melatonin syntheses and secretion explains by 68 % the differences observed in melatonin concentrations of digestive tract between controls and fasted quails. However 32 % of changes remains still unexplained by these sources. Thus it is possible that melatonin secreted in digestive system of quails is involved in signalling for energy balance.
Bubenik GA, Ball RO & Pang SF (1992) The effect of
food deprivation on brain and gastrointestinal tissue levels of tryptophan,
serotonin, 5-hydroxyindoleacetic acid, and melatonin. J. Pineal Res. 12:7-16.
Hohtola E, Hissa R, Pyörnilä A, Rintamäki
H & Saarela S (1991) Nocturnal hypothermia in fasting Japanese quail:
the effect of ambient temperature. Physiol. Behav. 49:563-567.
Rashotte ME, Basco PS, Saarela S and Henderson RP (1993)
Feeding schedule effects on the diurnal rhythms of heat production and
shivering thermogenesis in the pigeon. Society for Neuroscience Abstracts
19(1), 101.
Saarela S & Reiter R (1993) Function of melatonin
in thermoregulatory processes. Life Sci. 54:295-311.
ENERGY METABOLISM AND BODY TEMPERATURE IN THE BLUE-NAPED MOUSEBIRD (UROCOLIUS MACROURUS) DURING TORPOR
Schaub, R.& Prinzinger, R.
AK Stoffwechselphysiologie, Zoologisches Institut, Johann Wolfgang Goethe-Universität, Siesmayerstr. 70, D-60323 Frankfurt am Main, Germany
Mousebirds are a very old species of birds, endemic to Africa. They belong to an own order, the Coliiformes within they form with six species the one and only class, the Coliidae. Mousebird are herbivoruos, they prefer the arid and semi-arid thornbush and steppe areas, which on the one hand garantuees safety and on the other hand offers the basement for their special kind of food. They feed mainly on leafs, bud and fruits. This kind of food is low in energy, so the birds face relative fast a situation of energy deficiency To survive these times of starvation, they have developed a special physiological strategy which is called torpor, a state in which metabolic rate and body temperature are decreased to a very low level to save energy. Entrance into, maintenance of and arousal from torpor are active and regulated processes. First we sampled data about metabolic rate and body temperature under normal feeding conditions (food ad libitum), and then we rduced food successively to induce a state of energy deficiency which was followed by torpor in the night phases.
For the measurement of energy metabolism we used the method of indirect calorimetry. For this the oxygen consumption was measured in an open flow system. Body temperature was monitored parallel with a telemetric implant which garantuees monitoring without any external susceptible factors. With these two experimental setups it was possible to get informations continuously over long periods. So we can make statements about diurnal cycles in the monitored parameters.
Furthermore first results about the course of day of the plasma glucose level are shown. With these data we hope to get furher informations about the correlations between energy demand and energy preparation and the regulation during the state of torpor.
Supported by grants of the Deutsche Forschungsgemeinschaft DFG (Pr 202/4-1).
ENERGETICS AND BODY TEMPERATURE REGULATION IN PIGEONS AND DOVES OF EXTREME HABITATS
Schleucher, E.
AK Stoffwechselphysiologie, Zoologisches Institut, Johann Wolfgang Goethe-Universität, Siesmayerstr. 70, D-60323 Frankfurt, Germany
Endotherms adapt to their environment by varying their energy metabolism. However, only few comparative studies have examined the physiology of closelyrelated species living in different environments. The family Columbidae is ideal for such a study because its 300 species occur worldwide in almost every habitat except for the polar and alpine regions. Interestingly, only little is known about their strategies of adaptation to extreme climatic and nutritional conditions. In the present study basal metabolic rates (BMR) of 17 species were measured via O2consumption / CO2production, and compared with respect to taxonomy and biogeography. The tested bird species originated from a wide range of temperate, tropical, mesic and arid habitats. To account for differences in food utilization, fruit doves were incorporated in the study.
Previous allometric analyses predict low metabolic rates for pigeons compared with other birds, but the enlarged data set presented here indicates that their metabolism is intermediate on average. Lowered BMRs (- 20 %) were found only in specialists such as species confined to hot and arid regions, fruit doves and weak flyers.
Body temperature Tb was measured using telemetry in undisturbed birds in selected experiments. There was an apparent tendency to adaptations in Tb regulation to extreme environmental conditions. For instance, a well developed tolerance of elevated Tb (up to 45 °C without ill effects) was evident in the desert species. These physiological features enable them to be active even at high ambient temperatures (Ta). The fruit doves living on food of low nutritional quality had Tb below the expected values.
The ontogeny of pigeons, especially the development of thermoregulation, also plays an important role in the process of adaptive radiation of the columbids. The arid adapted Diamond Doves (body mass W = 38 g) and Namaqua Doves (W = 36 g) are typical opportunistic breeders. Young hatch at W of 1.9 - 2.0 g and are ectothermic. Because of rapid growth they achieve endothermy at an age of 5 days: individuals subjected to low Ta have higher metabolic rates than at Ta in the range of thermoneutrality (of adult birds). A high parental energetic effort is required during the phase of maximum growth while the total energetic cost per young raised is low because of the short development time and the small clutch size.
The thermoregulatory abilities and the energetics of Columbidae can be considered as main factors contributing to the species richness and successful adaptation of this family to a wide variety of habitats.
HEAT THERMOSENSITIVITY OF THE BRAIN AND SPINAL CORD IN GREENFINCHES
Sedunova, E.V.
Sechenov Institute of Evolutionary Physiology and Biochemistry, Russian Academy of Sciences, St.Petersburg, Russia
Heat thermosensitivity of 13 brain areas as well as of the thoracic and lumbar parts of the spinal cord was studied in free-moving greenfinches at the different ambient temperatures.
Clear vasomotor (sharp increase of the skin leg temperature) but not respiratory responses were observed only during local heating of the anterior hypothalamus (POA/POM) and the dorsal part of medulla. In contrast to ducks (Martin et al., 1981), there were no thermoregulatory reactions to heating of the midbrain in greenfinches. The minimum heating necessary to initiate a reaction in the thermoneutral zone was 0.7 to 1.0 °C (POA/POM) and 1.8 to 2.1 °C (medulla). The corresponding thresholds were 40.9 0.1 °C and 41.8 0.4 °C; and peak effector reaction was observed at initial brain temperatures of 39.8 to 40.2 °C.
Spinal heating led both to leg-temperature increase and polypnoea. The minimum heating for vasomotor reaction activation was 0.3 to 0.5 °C. During thoracic stimulation the vasomotor threshold was 41.7 0.2 °C, and the polypnoea threshold was 44.3 0.1 °C. Changes in the ambient temperature resulted in corresponding shifts of the thresholds. During lumbar stimulation the reaction thresholds were higher by 1.2 to 1.5 °C and the strength of the reactions was weaker; at low ambient temperature (14 to 17 °C) no polypnoea was observed. It is important to note that in lumbar heating the peak vasomotor reaction was observed if the initial spinal temperature was 42.0 to 43.5 °C, but not 41.2 to 42.2 °C as was the case with thoracic thermal stimulation. Also, at higher ambient temperatures lumbar heating appeared to be more effective. Therefore, in moderate hyperthermia when heat stimulation of the thoracic zone produced only weak effector response, lumbar heating maintained the ability to activate vasomotor reactions promoting an additional heat loss.
The data suggest that in passerines like in other birds the spinal cord
plays the leading role in central thermosensitivity. Probably the mammalian-like
hypothalamic thermosensors described in electrophysiological experiments
(Simon et al., 1977; Nakashima et al., 1987) participate in specific control
of brain temperature rather than in thermoregulation of the whole organism,
as it was proposed by Kuhnen & Jessen (1994) for goats.
Kuhnen, G. & Jessen, C. Am. J. Physiol. 267:
R355-R359 (1994)
Martin, R., Simon, E.& Simon-Oppermann, Ch. J.
Physiol. 314: 161-173 (1981)
Nakashima, T., Pierau, F.-K., Simon, E. & Hori, T.
Pfl.
Arch. 409: 236-243 (1987)
Simon, E., Hammel, H. & Oksche, A. J. Neurobiol.
8: 523-535 (1977)
CIRCADIAN PERSPECTIVE ON FEEDING, FASTING AND AMBIENT TEMPERATURE EFFECTS IN THE ZEBRA FINCH (TAENIOPYGIA GUTTATA)
Sedunova, E.V.1, Rashotte, M.E.2, Pastukhov, Iu. F.1, Pate, K.N.2 & Johnson, F.J.2
1Sechenov Institute of Evolutionary Physiology and Biochemistry, St. Petersburg, Russia
2Department of Psychology, Florida State University, Tallahassee, USA
The successful adaptation of the zebra finch to arid desert regions has encouraged study of its water economy and metabolism in response to variation in ambient temperature (Ta). This species, which withstands extended periods of water-deprivation, shows a linear increase in metabolic rate (MR) as Ta decreases from about 30 °C to 5 °C, even after 30 days without water (Cade, Tobin & Gold, 1965; Calder, 1964). The response of this species to an extended period of food-deprivation has not been described. The small size of this bird and the rapid passage of food through its digestive tract suggest that it could face a serious energetic challenge if food were withdrawn for more than a few hours (Cade, Tobin & Gold, 1965). We characterized the zebra finch's circadian rhythms in MR, RQ, subcutaneous back temperature (Ts), and locomotor activity (LA) during ad libitum feeding and during a period of food deprivation that stretched across almost two circadian cycles. In the feeding and fasted states, we also measured the zebra finch's response to short-term changes in Ta during the light and dark phases.
The finches were adapted to living individually in Ta = 26 °C and L:D 14:10 in a chamber that allowed flight activity (perches at each end). Mixed seeds and water were normally available in dispensors at floor-level in mid-cage. In ad lib feeding, there were pronounced day-night differences in all measures: MR (2.2 versus 1.2 ml/O2/min/bird), RQ (0.9 versus 0.7), Ts (36 °C versus 32.5 °C), LA (episiodic versus zero). Fasting had a large effect on day and night responses. In the first light phase without food, MR gradually decreased (from 2.2 to about 1 ml/O2/min/bird), RQ = 0.7, and LA became high and sustained. In the following dark phase, MR and Ts were exceptionally low (0.6 ml/O2/min/bird; 26.5 °C) and LA remained at zero as was usual for the dark. The next light phase began (after 34h without food) with high LA for about 1.5h before the birds became quiescent, sitting on the floor of the chamber with feathers erected; MR, RQ and Ts remained low. Food was returned part way through this light phase, after about 42h of fasting.
Responsiveness to variation in Ta was studied during approximately 7h "probe" periods when Ta decreased to 1 °C and then increased to 31°C (rate = 10 °C/h). A Ta probe was carried out during both the light- and dark-phases in ad lib feeding, and in the dark phase of the second fasting day when MR was exceptionally low. There was a linear relationship between MR and Ta in the range 1 °C to 31 °C in all probes. The birds remained behaviorally quiescent during the dark-phase cold probes.
The adult zebra finch, adapted to Ta = 26 °C, withstands an approximately 2-day fast by altering thermal, metabolic and locomotor activity in the light phase, and thermal and metabolic activity in the dark phase. We demonstrate that its response to ambient temperature probes involves metabolic, but not locomotor, responses, and that the metabolic response is comparable in light and dark phases, even in dark-phase fasting conditions when the birds are in a hypometabolic state.
Cade,T.J., Tobin, C.A. & Gold, A. Physiol. Zool.
38:9-33 (1965)
Calder, W.A. Physiol. Zool. 37:400-413 (1964)
DEVELOPMENT OF AVIAN THERMOREGULATORY CENTRAL NERVOUS MECHANISMS DURING THE PERINATAL PERIOD
Tzschentke, B., Basta, D. & Nichelmann, M.
Institut für Biologie, Humboldt-Universität zu Berlin, Germany
The aim of the study was to investigate the development of thermosensitivity of the preoptic area of the anterior hypothalamus (PO/AH) during the perinatal period, the influence of different prenatal temperature experiences on the postnatal hypothalamic thermosensitivity and the development of the thermoregulatory set point in birds.
The experiments were carried out in embryos (28th and 33th day of incubation) and 1-, 5- and 10-d-old ducklings of the Muscovy duck (Cairina moschata) incubated at 37.5°C and at 35 and 38.5°C. Using the method of extracellular recordings thermosensitivity of PO/AH-neurons after sinusoidal temperatur changes (40°C3°C) in brain slices was investigated. In birds incubated at 37.5°C the influence of bombesin (bolus injection 1µg/0,1 ml artificial cerebrospinal fluid) on hypothalamic thermosensitivity was tested. The thermoregulatory set-point was calculated by mesurement of colonic temperature.
The results can be summarized as follows:
Development of PO/AH thermosensitivity
(1) In contrast to adult birds and mammals (cold-sensitive (cs) neurons 10%) during the perinatal period in embryos and 1- to 5-d-old Muscovy ducklings the percentage of cs- PO/AH-neurons was much higher (30%) and decreased to 14% until to the 10th day post-hatching. It suggests a possibly thermoregulatory role of cs-PO/AH-neurons in prenatal and juvenil birds.
(2) Temperature guardian neurons, which were sensitive exclusively in a small temperature range not more than 0.4°C around 36 and 42°C (ducklings) and 38 and 40.5°C (embryos) were found. Possibly, they react to signal extreme brain temperatures and play a role in the wide band control (Bligh, 1966) of body temperature.
(3) Bombesin-application induced an alteration of the hypothalamic thermosensitivity in embryos and in ducklings similar to adult mammals (Schmid et al., 1993).
Influence of prenatal temperature experience on postnatal hypothalamic thermosensitivity
Proportion of temperature-insensitive and warm- and cold-sensitive neurons in 10-d-old Muscovy ducklings
Prenatal temperature experience to higher or lower incubation temperatures than 37.5°C induced a clear change in the thermosensitivity of PO/AH-neurons (Fig.). The increase in the proportion of cs-neurons and also a decline in the number of warm-sensitive PO/AH-neurons of warm incubated ducks indicates a decrease and the reduction in the number of cs-neurons in cold-incubated ducks indicates an increase in the total warm-sensitivity in the PO/AH.
In 1- and 5-d-old Muscovy ducklings the change of the PO/AH thermosensitivity after prenatal cold- or warm-load was unspecific.
Development of thermoregulatory set point
The thermoregulatory set-point increased from day 1 untill day 10 and was influenced by different prenatal temperature load.
Bligh, J. Biol. Rev. 41: 317-367 (1966)
Schmid, H., Jansky, L., Pierau, Fr.-K. Am. J. Physiol.
264: R449-R455 (1993)
Supported by the Deutsche Forschungsgemeinschaft (Tz 6/2-1).
<TOP> <HOME> <Esa Hohtola's home page>